

400-820-8531




背景介绍
有些生物虽在千万年前分道扬镳,却仍暗藏“藕断丝连”的进化密码——它们形态相近、核心发育程序稳定,却在分子层面悄然分化,成为解锁“进化如何平衡保守与创新”的关键线索。
美国宾夕法尼亚大学John I. Murray团队正是瞄准这一科学谜题,在Science发表题为“Lineage-resolved analysis of embryonic gene expression evolution in C. elegans and C. briggsae”的研究,以秀丽隐杆线虫(C. elegans)和布氏隐杆线虫(C. briggsae)为模型,系统性揭开了基因表达进化的深层规律。这两种线虫早在 2000 万年前从共同祖先分化,形态近乎一致,胚胎发育细胞谱系高度保守,堪称破解基因表达进化的“天然理想模型”。
或许有人会疑惑:研究线虫进化,其科研价值究竟体现在何处?事实上,线虫与人类共享大量同源基因,其基因调控网络宛如“简化版人类发育模型”。该研究不仅助力我们厘清“生物如何在维持核心功能的同时适应环境”——即进化是“修修补补”而非“推倒重来”的内在逻辑,更为人类发育疾病研究提供了关键参考。许多人类发育异常的本质是基因调控网络紊乱,而线虫研究能帮助我们锁定这些网络的“关键节点”,解析其进化规律,进而更精准地破解人类疾病密码。
BIOPROFILE
17万+单细胞测序
揭开两物种的“基因默契”
研究团队借助10x Genomics平台,对布氏隐杆线虫(C. briggsae)胚胎开展单细胞 RNA 测序,样本覆盖原肠胚形成期至终末分化的完整发育阶段。经严格质控后,最终获得 178,818 个高质量细胞。将该数据集与已发表的秀丽隐杆线虫(C. elegans)单细胞胚胎图谱进行跨物种比对,结果显示,两种线虫的细胞在联合转录空间中聚类效果优异,可精准匹配共享细胞类型。进一步分析同源基因表达模式发现,跨物种标记基因的表达特征高度保守。这一结果证实,两种线虫不仅具备保守的胚胎发育谱系,更存在可稳定识别的同源分子特征,为深入开展发育过程中同源细胞与基因的比较研究奠定了坚实基础。
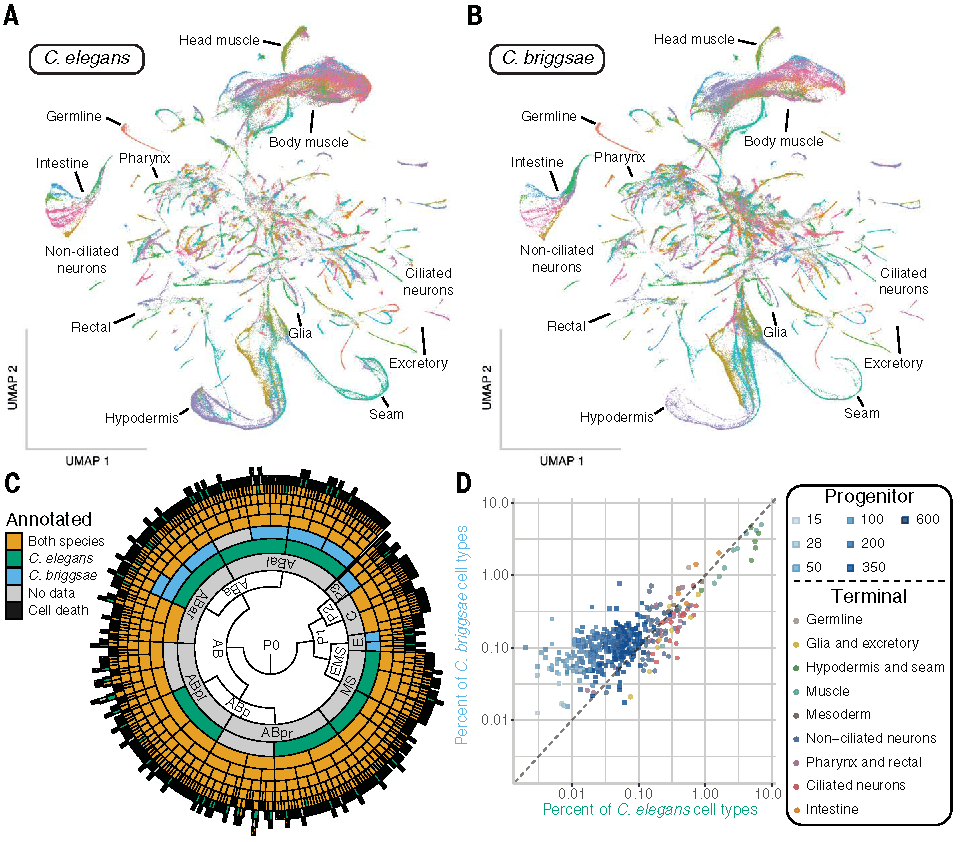 点击添加图片描述(最多60个字)
点击添加图片描述(最多60个字)
Fig1.线虫细胞类型注释中的同源性
进化不只有“传承”
更有“专属定制”
为了量化基因表达的进化程度,研究者使用Jensen-Shannon距离(JSDgene),基于所有同源细胞类型的表达水平[每百万转录本数(TPM)]进行计算。结果显示,5500个基因(约 55%)的表达模式高度保守(JSDgene<0.45),同时有1849个基因(约 18.5%)表现出显著表达分歧(JSDgene>0.65),且这种差异具有明确的功能倾向性。
进一步分析基因类型与表达进化的关联发现:广泛表达的基因通常高度保守,例如与细胞周期、发育相关的基因及转录因子;而特异性表达的基因更易呈现差异表达模式,如核激素受体(NHR)家族、化学受体相关的 G 蛋白偶联受体(GPCR)、七跨膜(7TM)受体家族以及神经肽。此外,在特异性或模式化表达的基因中,表达水平较高的基因更可能保持保守特性。
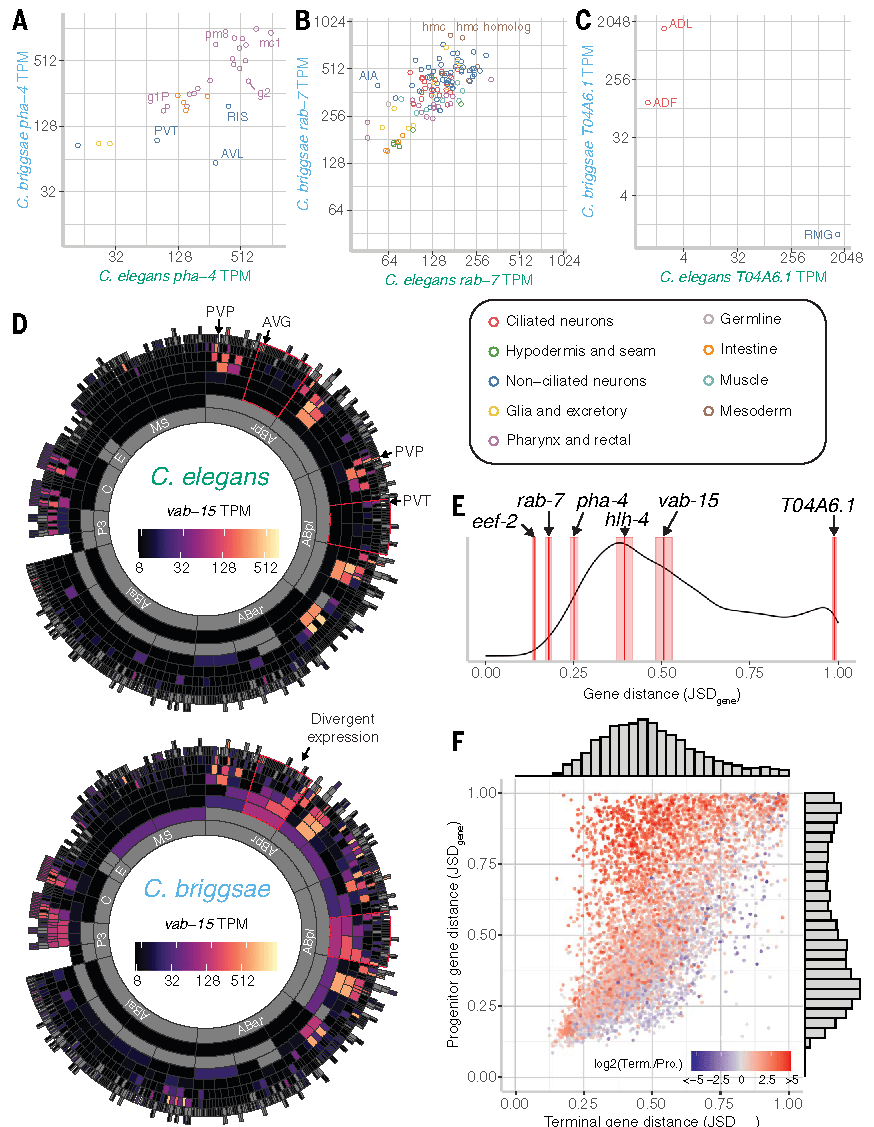 点击添加图片描述(最多60个字)
点击添加图片描述(最多60个字)
Fig2.直系同源基因在胚胎发育过程中的基因表达模式
基因表达的
“广度约束”与“时序灵活”
研究通过计算Tau值量化基因表达广度(0 代表广泛表达,1 代表细胞特异性表达),发现基因的表达广度在两物种间高度一致(R2=0.77)。这表明,基因在“广泛表达”与“细胞特异性表达”之间的转换受到严格的进化约束。
与之形成鲜明对比的是,基因表达的时间动态具有显著灵活性——存在表达起始时间或持续时间的物种特异性变化。例如,秀丽隐杆线虫中生殖细胞相关基因gld-1和cep-1的转录本在合子系中快速消失,而在布氏隐杆线虫中可持续至终末体细胞阶段。这种“广度保守、时序灵活”的特征,揭示了基因表达进化的差异化调控策略。
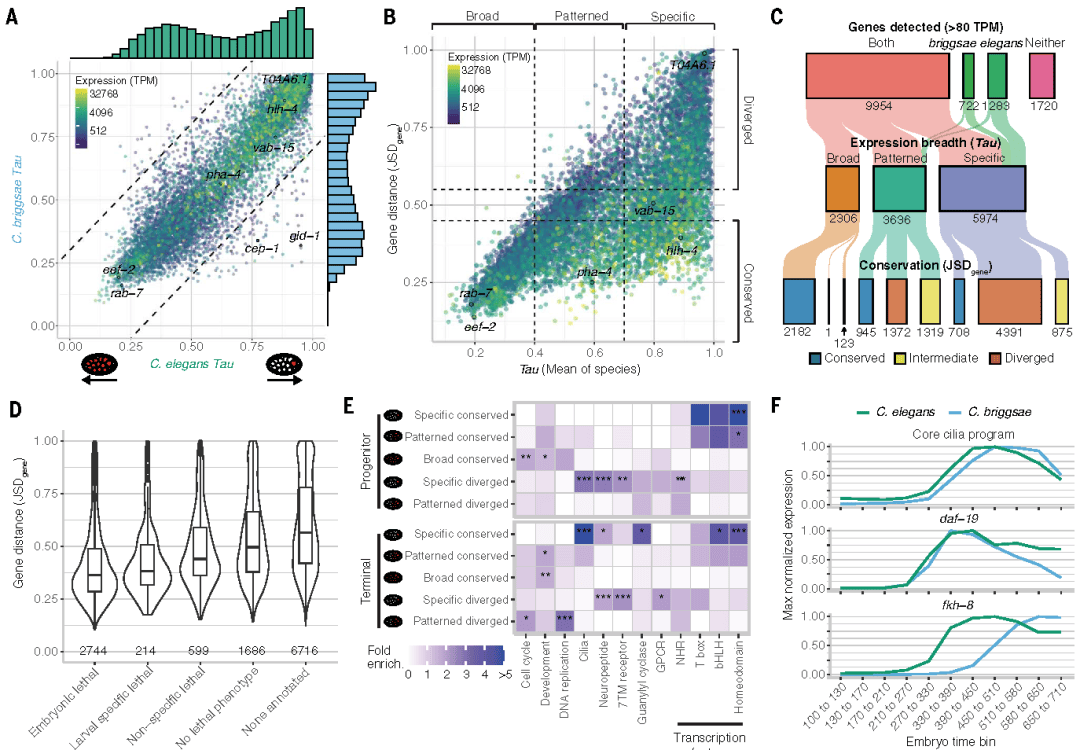 点击添加图片描述(最多60个字)
点击添加图片描述(最多60个字)
Fig3.基因表达广度与保守性的交集揭示了基因家族水平的差异
进化的“沙漏密码”
胚胎中期为保守峰值
研究团队利用单细胞转录数据分析各细胞类型的表达谱变化,通过计算细胞类型间的Jensen-Shannon距离发现,胚胎发育中期(约200 细胞阶段)的细胞转录组相似性最高,细胞距离达到最小值,随后在晚期祖细胞和终末细胞中逐渐上升。这一结果与经典的“发育沙漏模型”高度契合。
值得注意的是,不同细胞谱系的保守模式存在异质性:例如通向ASG神经元的谱系在200细胞阶段相似度最高,而头部体壁肌肉谱系的转录组相似性随发育进程持续升高,提示发育约束存在组织特异性差异。
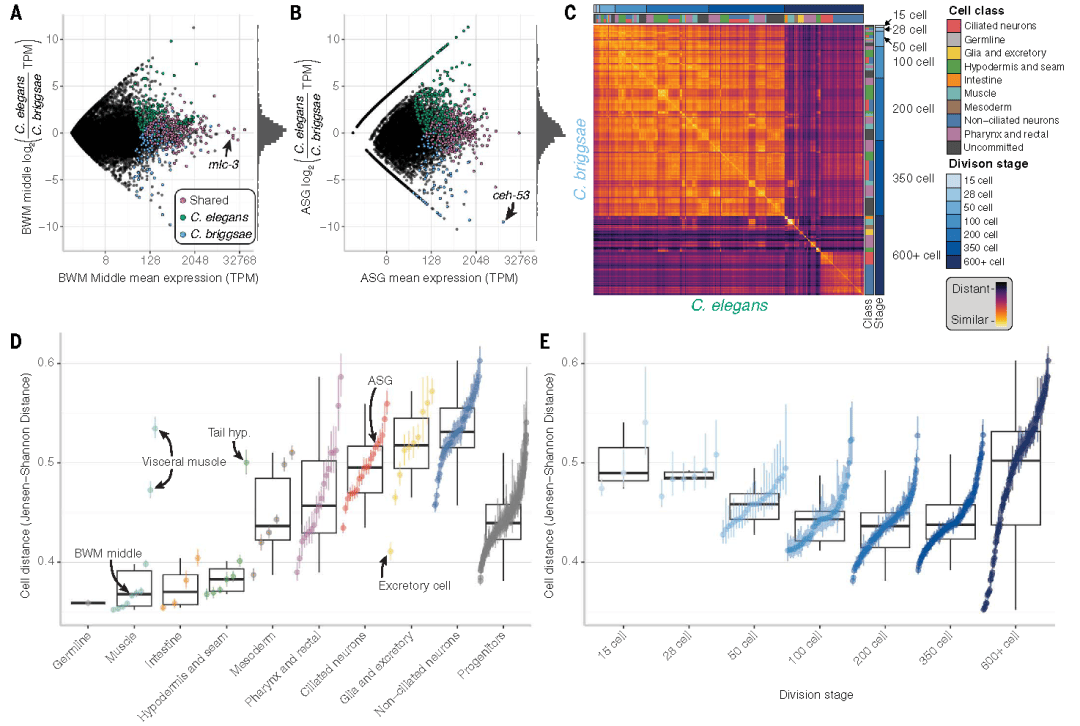
Fig4.细胞功能决定同源细胞类型之间转录组的差异
细胞类型特异性表达
核心功能保守与特化功能分化
细胞距离反映了一种细胞类型转录组谱的整体相似性,涵盖广泛表达基因与决定细胞命运、特化功能的细胞类型特异性基因之间的差异。基于此,研究团队鉴定并比较了两种物种中每种细胞类型的特异性标记基因。结果显示,从细胞类型来看,神经元和排泄细胞的转录组分化程度最高,而肌肉、肠道等执行基础生理功能的细胞类型则更为保守。这一现象提示,细胞类型的功能需求是驱动基因表达分化的关键因素。
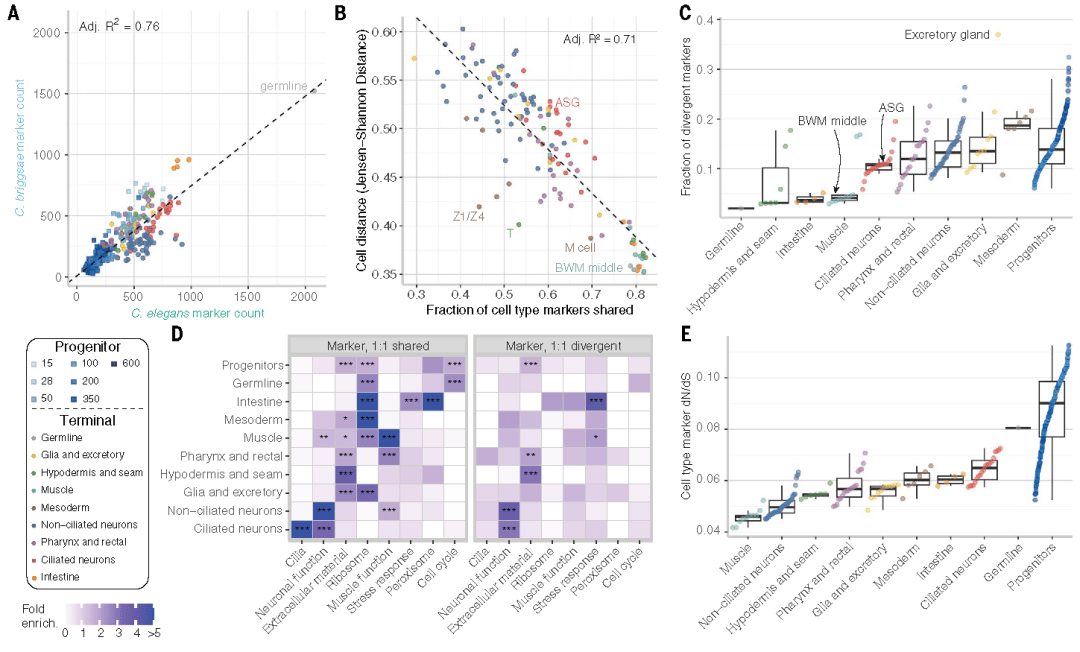
Fig5.细胞类型特异性的基因表达突显了细胞类型进化的复杂性
基因拷贝驱动表达多样化
呈现“退化为主、创新为辅”特征
基因会经历复制与丢失过程,有时会发生新功能化或部分功能化,进而承担新的生物学角色。研究发现,细胞类型标记基因富集于1:1直系同源基因,但不同细胞类型在两物种中标记基因的总数及复杂同源关系(如1:多、多:多)标记基因的比例存在显著差异,这种差异既与发育阶段相关,也与细胞类别相关。
对非1:1同源基因(尤其是1:2拷贝关系基因)的分析显示,基因拷贝数是塑造表达模式进化的重要力量。多数基因拷贝趋向功能退化,仅少数拷贝通过新功能化获得独特表达模式。例如,AP-2 转录因子家族中,部分旁系同源基因在保留祖先表达特征的同时,获得了生殖系或早期胚胎特异性表达,为物种适应性进化提供了分子基础。
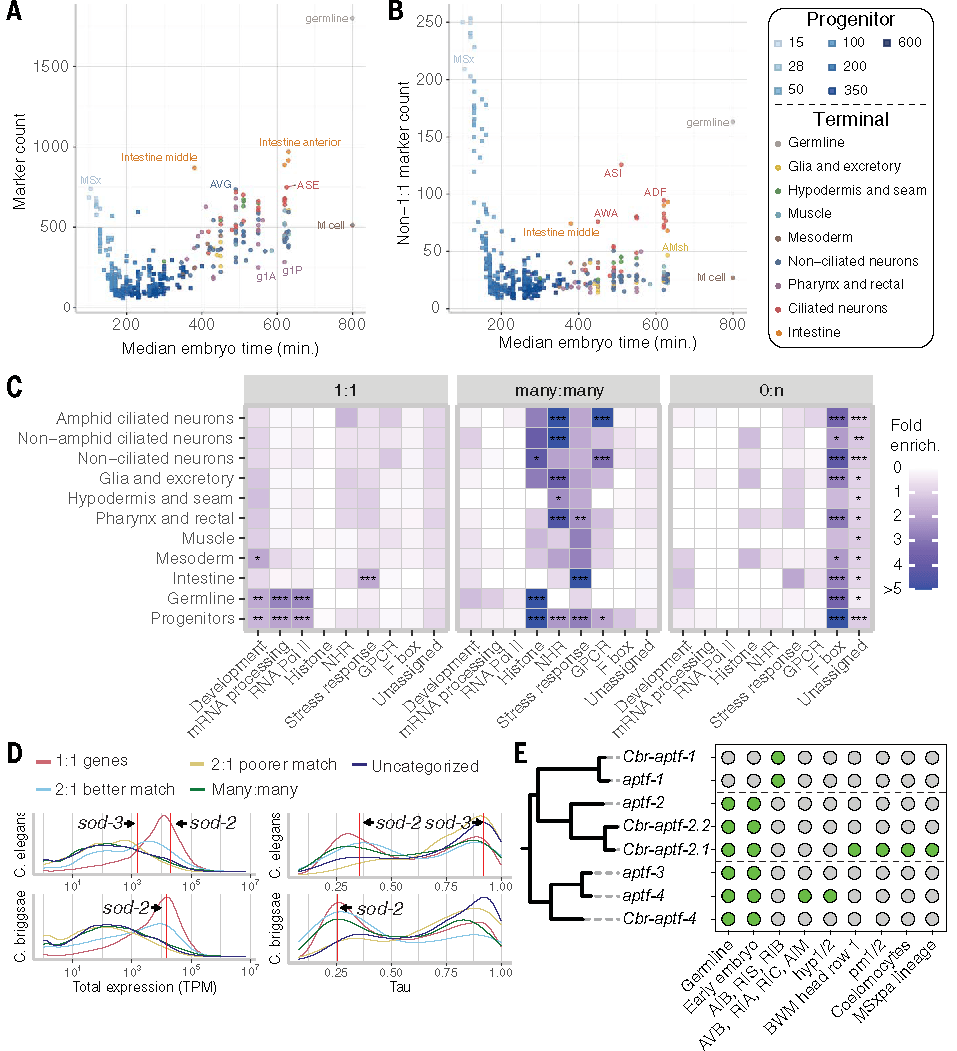 点击添加图片描述(最多60个字)
点击添加图片描述(最多60个字)
Fig6.非1:1直系同源关系的基因的表达与保守性
文章小结
从2000万年前的共同祖先,到如今的“同源不同貌”,这两种线虫用自己的基因密码,写下了进化的智慧:坚守核心,灵活应变。而科学家通过单细胞测序技术,读懂了这份跨越千万年的“进化日记”,也为我们打开了理解生命多样性的新窗口。生命的进化从来不是孤勇前行,而是在传承与创新中寻找平衡,这大概就是自然最神奇的智慧吧!
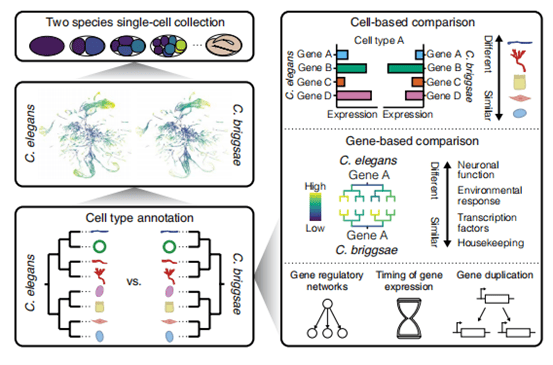 点击添加图片描述(最多60个字)
点击添加图片描述(最多60个字)
Fig7.两种线虫物种的发育细胞图谱
拜谱小结
拜谱生物深度布局单细胞研究领域,可提供全面的单细胞转录组测序与分析服务,同时涵盖蛋白质组学、修饰组学等核心组学技术,能通过多组学数据整合挖掘与深度机制解析,精准匹配演化生物学、发育生物学等领域的研究需求。从技术支持到数据分析全链条赋能,助力科研人员解锁基因表达进化等关键科学问题,冲刺高分文章!欢迎咨询合作,共同推进科研突破!
参考文献
Large CRL, Khanal R, Hillier L, Huynh C, Kubo C, Kim J, Waterston RH, Murray JI. Lineage-resolved analysis of embryonic gene expression evolution in C. elegans and C. briggsae. Science. 2025 Jun 19;388(6753):eadu8249.





关注公众号





关注小红书


