

400-820-8531




光照和温度是影响植物生长发育的两种重要的环境因素,它们不仅影响植物的形态建成,还调控着植物对环境的适应性。ELONGATED HYPOCOTOYL5(HY5)和PHYTOCHROME INTERACTING FACTORs(PIFs)是两种重要的植物生长的光调控因子,它们之间的相互作用尚不明确。
2024年8月由中国浙江大学周艳虹教授团队在知名期刊PNAS发表题为“SlCPK27 cross-links SlHY5 and SlPIF4 in brassinosteroid-dependent photo- and thermo-morphogenesis in tomato”文章,研究发现,激活的番茄HY5(SlHY5)会触发一个钙依赖蛋白激酶SlCPK27的转录。SlCPK27与SlPIF4相互作用,并在Ser-252和Ser-308磷酸化位点上对其进行磷酸化,以促进其降解。SlPIF4主要通过激活油菜素内酯(BR)生物合成中的一个关键基因SlDWF的转录来促进下胚轴的伸长。这种SlHY5-SlCPK27-SlPIF4-BR级联反应不仅在光形态建成中起着至关重要的作用,还调节热形态建成。拜谱生物为该研究提供了RNA-seq技术服务。

英文标题:SlCPK27 cross-links SlHY5 and SlPIF4 in brassinosteroid-dependent photo- and thermo-morphogenesis in tomato(PNAS,2024.08)
中文标题:SlCPK27在番茄中通过油菜素内酯依赖的光和热形态建成中交叉连接SlHY5和SlPIF4
客户单位:浙江大学
研究材料:番茄幼苗
拜谱提供技术:RNA-seq
技术路线:
一、研究结果
01. SlCPK27基因的发现和功能确定
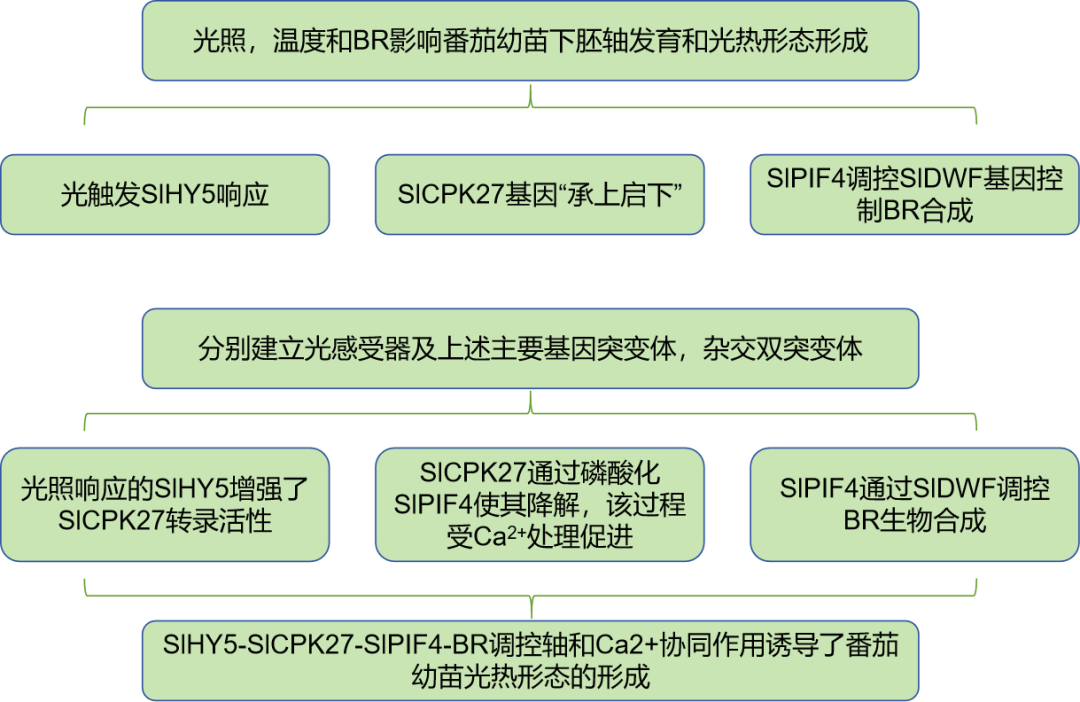
前期研究通过RT-qPCR发现SlCPK27表达量在白光(W)、蓝色(B)、红色(R)和远红色(FR)照射下均出现了显著的上升,光感受器突变植株与WT相比,该基因的上升幅度显著下降或不出现上升,说明该基因与植株光感是紧密相关的(图1 A~C)。
为了研究SlCPK27是否在光形态建成中起作用,在黑暗和光照条件下培养了两个纯合子系sicpk27 #2和sicpk27 #6。SlCPK27突变体在黑暗条件下的下胚轴长度与WT相似,但在光照条件下的下胚轴长度明显长于WT。
02.SlCPK27基因与HY5相互作用研究
针对光形态建成重要调节因子HY5的研究中发现,其具有与SlCPK27相似的光响应趋势,在HY5突变植株slhy5 #1和slhy5 - oe的研究中发现,后者植株的下胚轴明显短于WT和slhy5 #1(图2),并发现slhy5 #1突变体中SlCPK27的光诱导表达受到抑制,但与WT相比,slhy5 - oe幼苗中SlCPK27的光诱导表达显著增加(图1D),并进行了电泳迁移转移实验(EMSA),发现his标记的SlHY5蛋白可以与SlCPK27的启动子结合(图1E)。
后续使用双荧光素酶(LUC)报告系统在烟叶中进行了瞬时表达实验,结果显示SlHY5显著增强了SlCPK27的启动子活性(图1G)。
为了验证SlCPK27和SlHY5之间的遗传相互作用,培育了对应的双突变体,发现其下胚轴长度与SlCPK27突变体接近(图1H)。
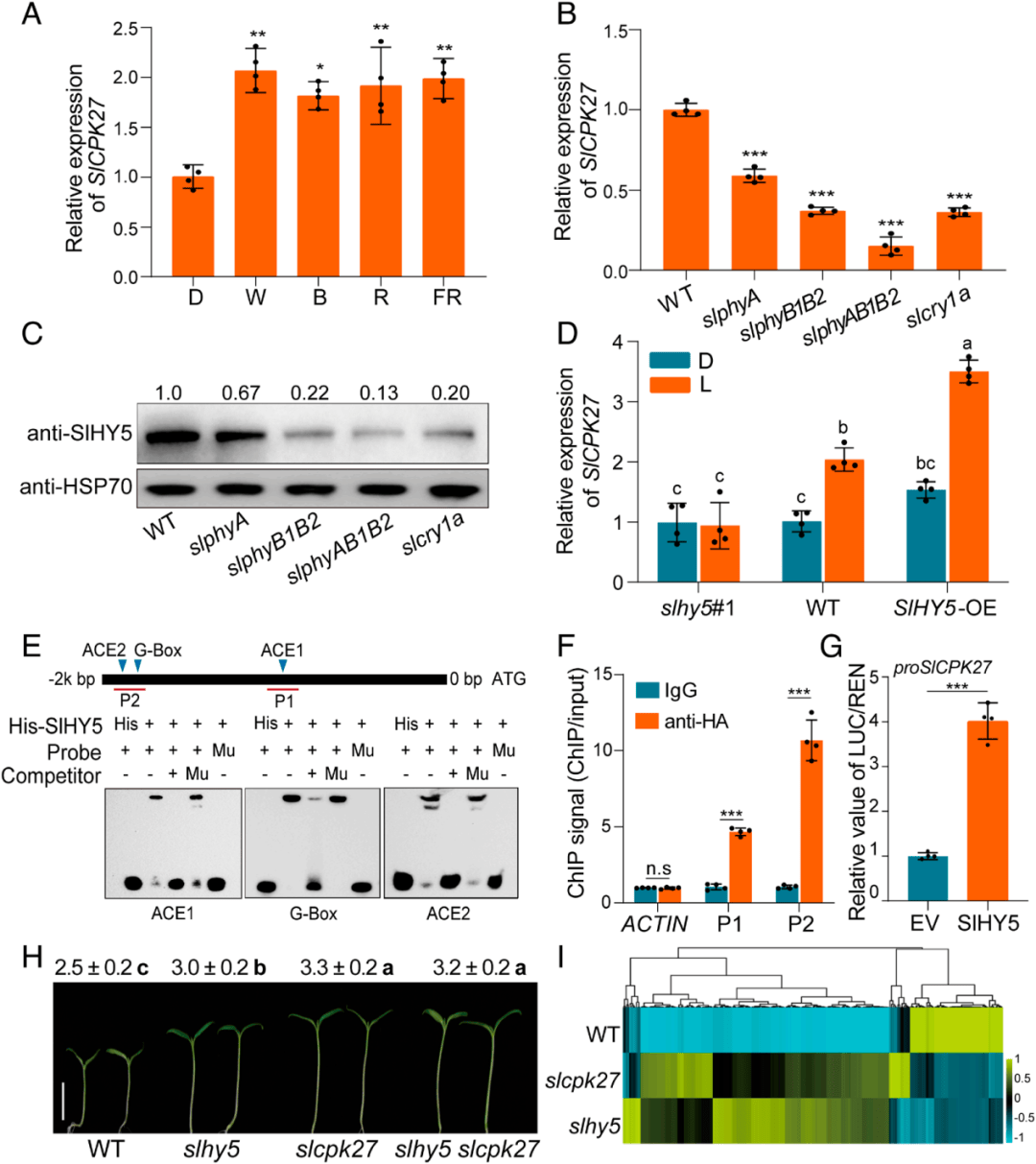 图1 SlCPK2与SIHY5相互作用(图源:Zhu C,Hu Z,Hu C, et al.,Proc Natl Aca
图1 SlCPK2与SIHY5相互作用(图源:Zhu C,Hu Z,Hu C, et al.,Proc Natl Aca
 图2 SIHY5突变植株HY5表达与植株萌发情况
图2 SIHY5突变植株HY5表达与植株萌发情况
(图源:Zhu C,Hu Z,Hu C, et al.,Proc Natl Acad Sci U S A. 2024)
03.SlCPK27基因与SlPIF4相互作用研究
为了解决slhy5调控SlCPK27如何抑制下胚轴伸长的问题,进一步研究SlCPK27和SlHY5之间的相互作用,作者对WT、SlHY5和SlCPK27幼苗在W光下生长4 d进行了转录组学分析。比值≥2或≤0.5,p值< 0.05的转录本被鉴定为差异表达基因(DEG)。基于这一标准,作者鉴定了416个SlCPK27调控的DEG和890个SlHY5调控的DEG(图3)。
作者通过酵母双杂交(Y2H)筛选来确定SlCPK27的潜在相互作用伙伴,在90个候选物中重点研究了与光信号通路相关的转录因子PIF4,酵母双杂交实验证实了SlCPK27和SlPIF4之间的相互作用(图4A),后使用Pull-down,双分子荧光互补(BiFC)实验验证了二者的互作关系,并在Ca2+处理后增强(图4D)。
在植物体内研究中发现,在slcpk27突变体在光照下的SlPIF4蛋白在核、质积累要比WT高得多(图4E,F),SlPIF4的光诱导降解被26S蛋白酶体抑制剂MG132抑制,这表明SlCPK27通过26S蛋白酶体途径促进了SlPIF4的光诱导降解(图4G)。
通过生成slpif4突变株和slcpk27/slpif4双突变株,发现二者相较于WT和slcpk27突变株,均表现出了短下胚轴的性状(图4H)。
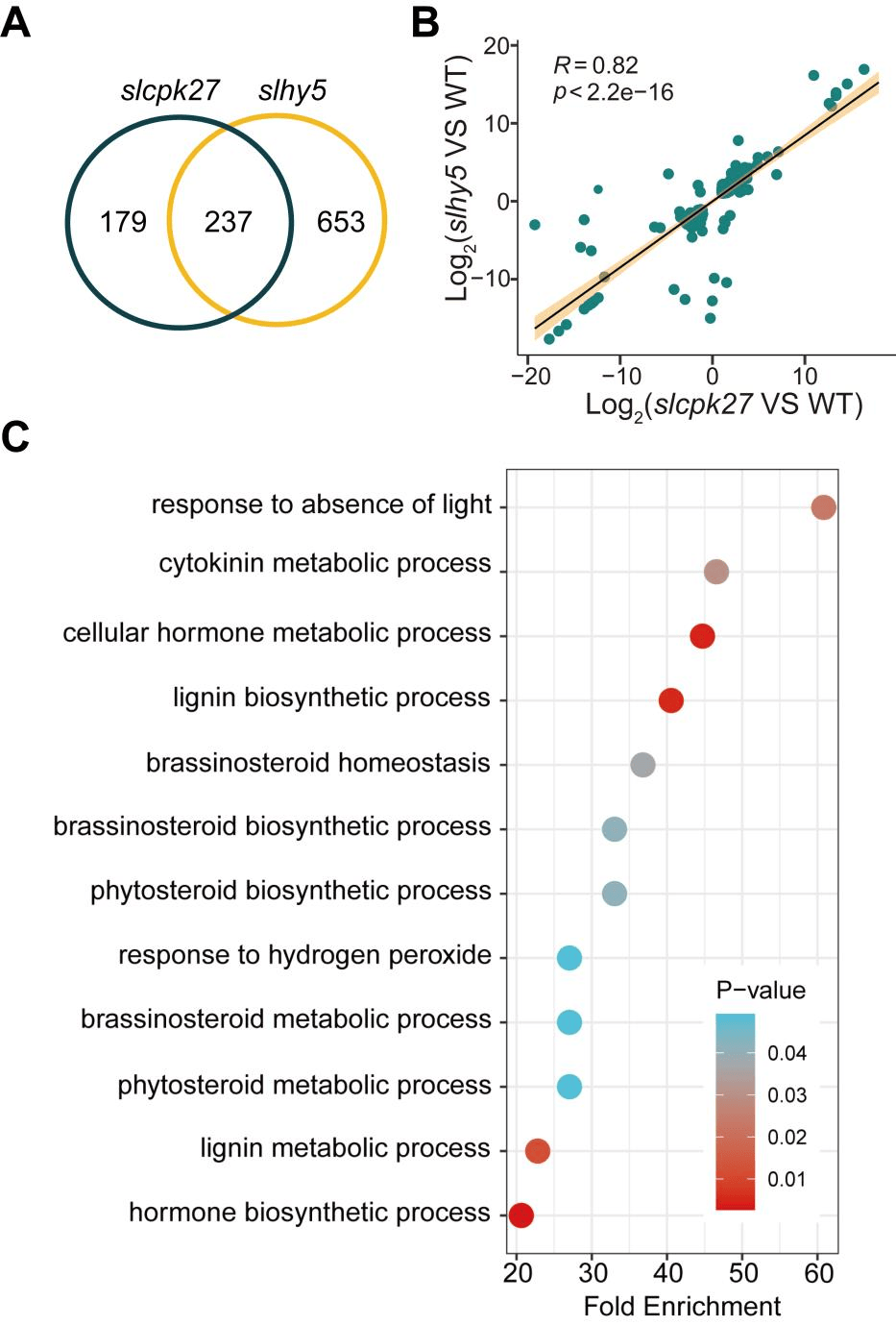 图3 SlCPK27和SlHY5共调控基因的转录组分析
图3 SlCPK27和SlHY5共调控基因的转录组分析
(图源:Zhu C,Hu Z,Hu C, et al.,Proc Natl Acad Sci U S A. 2024)
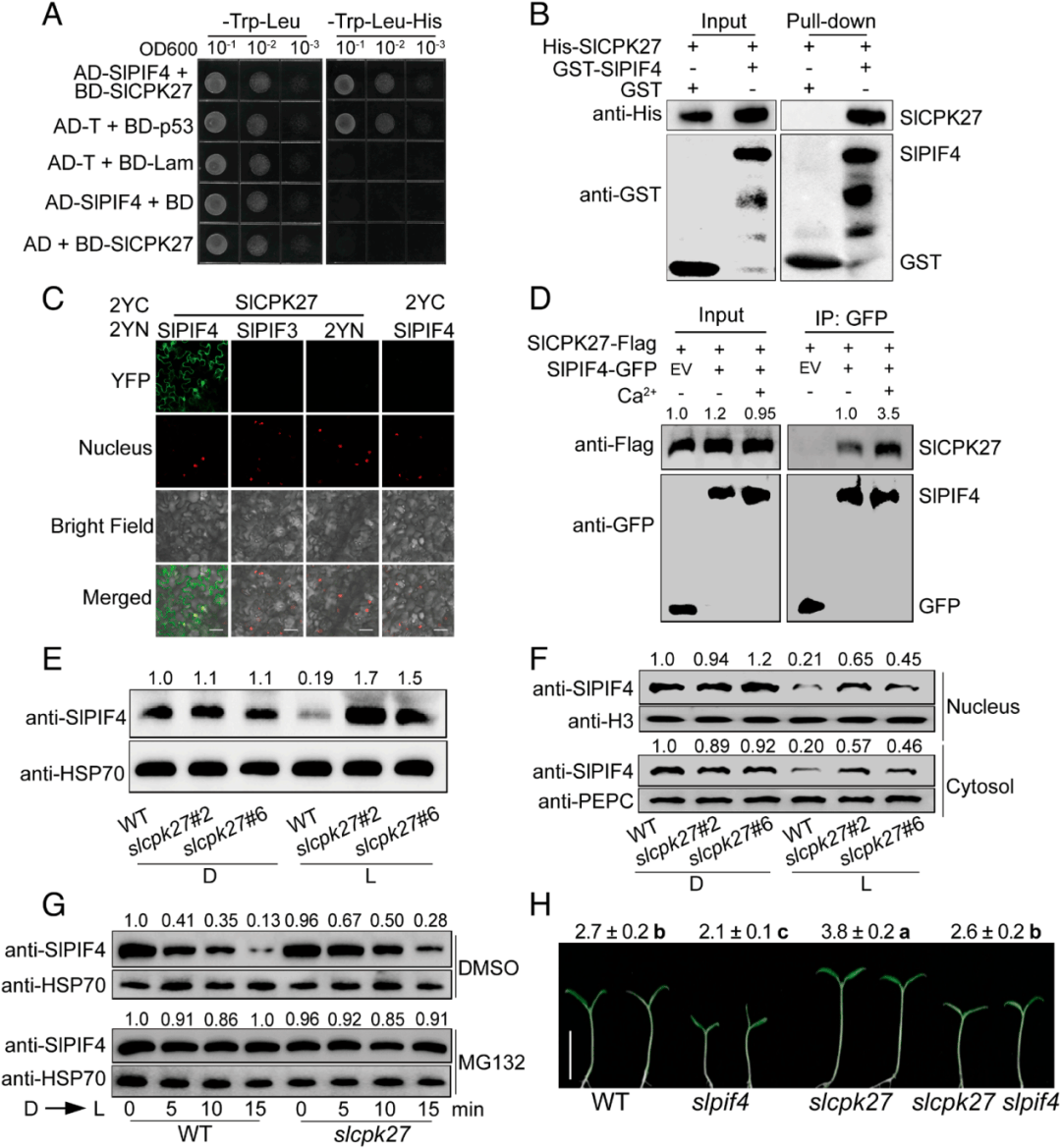 图4.SlCPK27与SlPIF4相互作用
图4.SlCPK27与SlPIF4相互作用
(图源:Zhu C,Hu Z,Hu C, et al.,Proc Natl Acad Sci U S A. 2024)
PIF蛋白磷酸化是光诱导PIF蛋白降解的先决条件,考虑到光触发Ca2+流入细胞质以促进植物的光形态建成,使用黄化苗进行CaCl2处理和H2O对照探究了Ca2+对该过程的影响,发现CaCl2处理增强了光诱导SlPIF4磷酸化,但EGTA和LaCl3处理抑制了SlPIF4磷酸化;考虑到cpk作为Ca2+信号解译器,作者随后通过使用体外磷酸化反应后纯化的大肠杆菌His-SlCPK27和MBPSlPIF4蛋白进行phos标签迁移转移实验,探索SlCPK27是否直接磷酸化SlPIF4。在His-SlCPK27存在的情况下,观察到MBP-SlPIF4的迁移率移位带,表明SlPIF4在体外被SlCPK27磷酸化(图5A)
Ca2+和光刺激进一步促进了这种磷酸化(图5B,C),同时,使用抗pSer /pThr抗体进行免疫印迹分析显示,SlCPK27-GFP磷酸化水平和激酶活性在光照后5分钟内增加(图5E)。这些数据共同揭示了SlCPK27以Ca2+和光依赖的方式磷酸化SlPIF4。
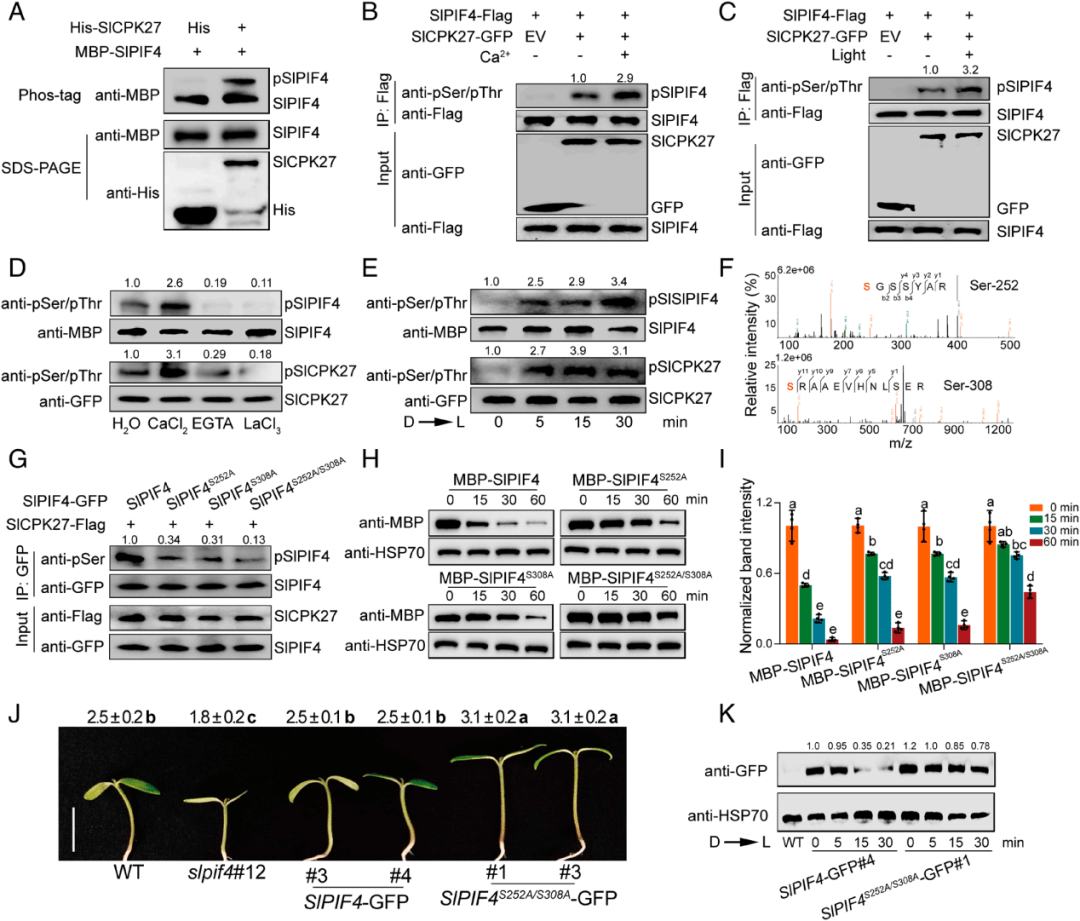 图5 SlCPK27磷酸化SlPIF4有助于光诱导SlPIF4蛋白降解
图5 SlCPK27磷酸化SlPIF4有助于光诱导SlPIF4蛋白降解
(图源:Zhu C,Hu Z,Hu C, et al.,Proc Natl Acad Sci U S A. 2024)
04.油菜素内酯(BR)在SlCPK27调控光形态形成中的作用
生长素和BR在pif4调控的下胚轴伸长中起关键作用,slpif4突变体在光照条件下,CS、BL和吲哚-3-乙酸(IAA)含量明显低于WT(图6A-C),使用24-表油菜素内酯(eBL)和合成生长素picloram (Pic)来区分BR和生长素在slpif4调控的下胚轴伸长中的作用。eBL显著促进了WT的下胚轴伸长,Pic仅部分挽救了slpif4的下胚轴缺陷表型(图7)。
转录组和RT-pcr结果显示slpif4突变体中SlDWF的转录物积累量减少,但与WT相比,其他BR生物合成基因的转录物积累量没有明显变化(图6D)。为了验证SlPIF4是否直接调控SlDWF的表达,作者进行了EMSA,发现MBP-SlPIF4可以结合SlDWF的启动子在所有g -box样基序上(图6E),并使用ChIP进一步进行了验证(图6F)。
在突变植株研究中发现,sldwf突变体的下胚轴较短,而SlDWF过表达(sldwf - oe)系的下胚轴较长。重要的是,slcpk27突变体(slcpk27 SlDWF)中SlDWF的突变破坏了slcpk27的长下胚轴表型,而slpif4中SlDWF的过表达恢复了slpif4的短下胚轴表型(图6H)。
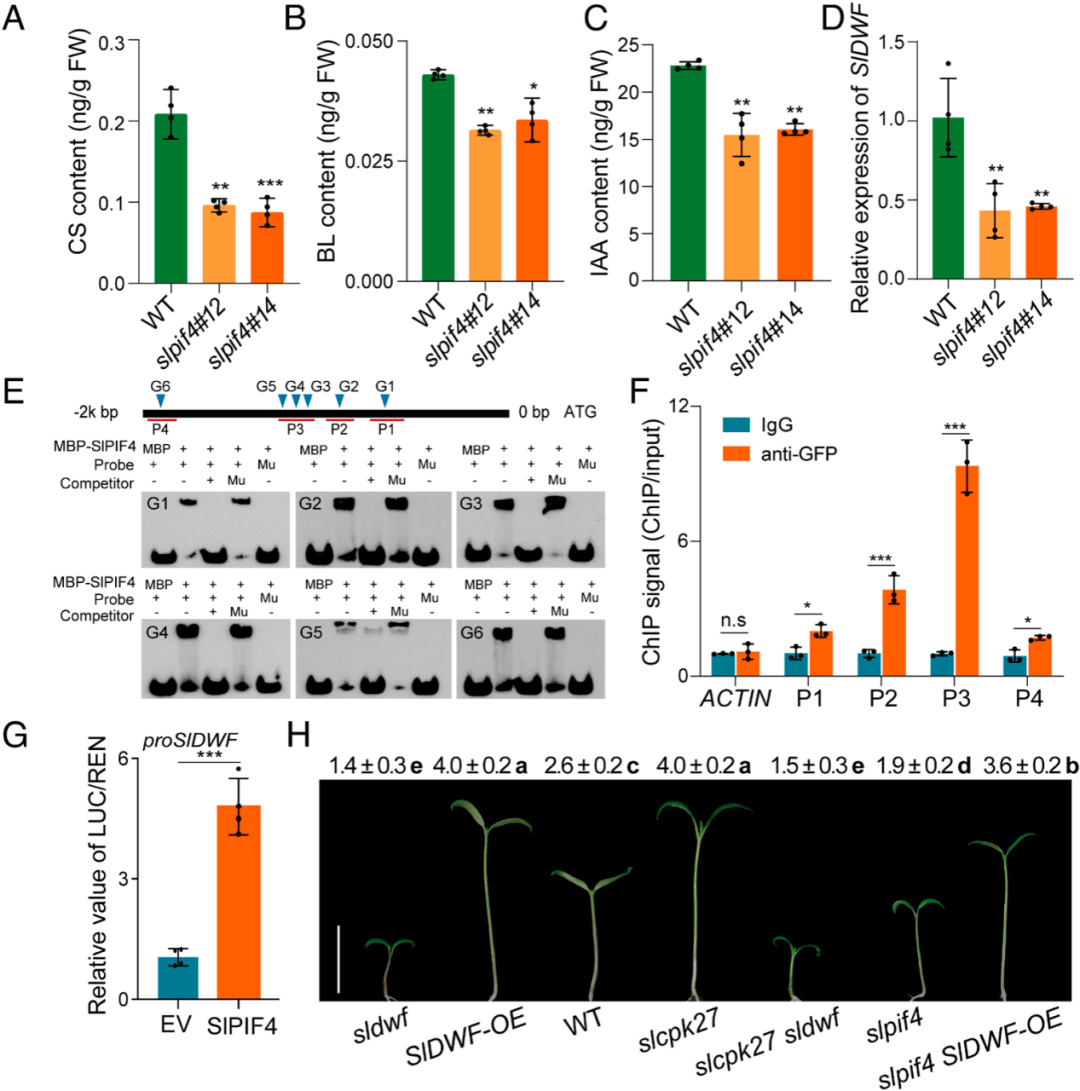 图6 SlPIF4通过触发BR生物合成负调控光形态建成
图6 SlPIF4通过触发BR生物合成负调控光形态建成
(图源:Zhu C,Hu Z,Hu C, et al.,Proc Natl Acad Sci U S A. 2024)
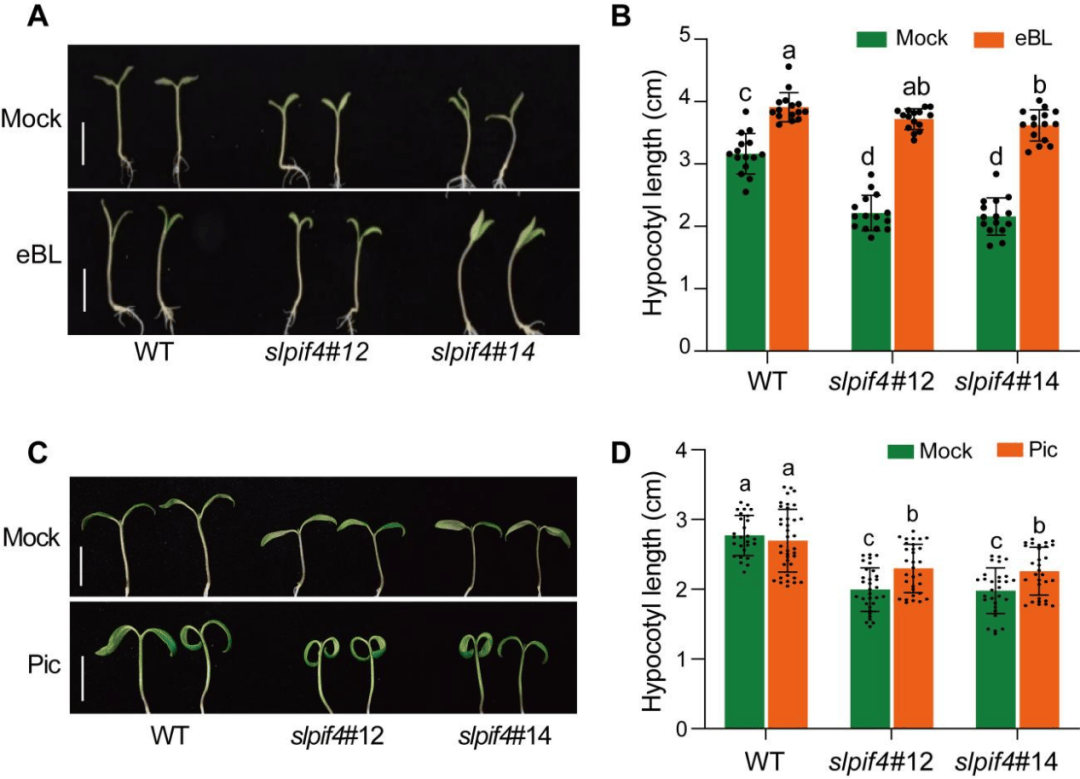 图7 BR和生长素在slpif4介导的下胚轴生长中的作用
图7 BR和生长素在slpif4介导的下胚轴生长中的作用
(图源:Zhu C,Hu Z,Hu C, et al.,Proc Natl Acad Sci U S A. 2024)
05.SlHY5-SlCPK27-SlPIF4-BR模块在热形态建成中的作用
除了光,温度也是调节植物生长发育的关键环境线索,为了确定SlHY5-SlCPK27-SlPIF4模块是否也参与热形态建成的调控,将光感受器突变体(slphyA、slphyB1B2、slphyAB1B2和slcry1a)在25℃或32℃的光照下培养。发现温暖环境下WT下胚轴显著增长,但是这一现象在突变体中均减弱或消失(图8A,B),此外,温暖的温度降低了SlHY5蛋白的积累,但增加了SlPIF4蛋白在WT幼苗中的积累(图8C),温暖温度显著降低了SlCPK27的转录量,增加了SlDWF的转录量,上述变化在突变体中也有所减弱,证实了以上光感受器和SlHY5-SlCPK27-SlPIF4-BR通路在植株热形态调控中的作用。
虽然温度显著影响了WT下胚轴长度,但对slhy5和slcpk27突变体的下胚轴长度影响甚微,而对slpif4和sldwf突变体的下胚轴长度没有显著影响(图8F,G),这一现象强调了SlHY5-SlCPK27-SlPIF4模块通过调节BR生物合成在热形态建成中的重要作用。
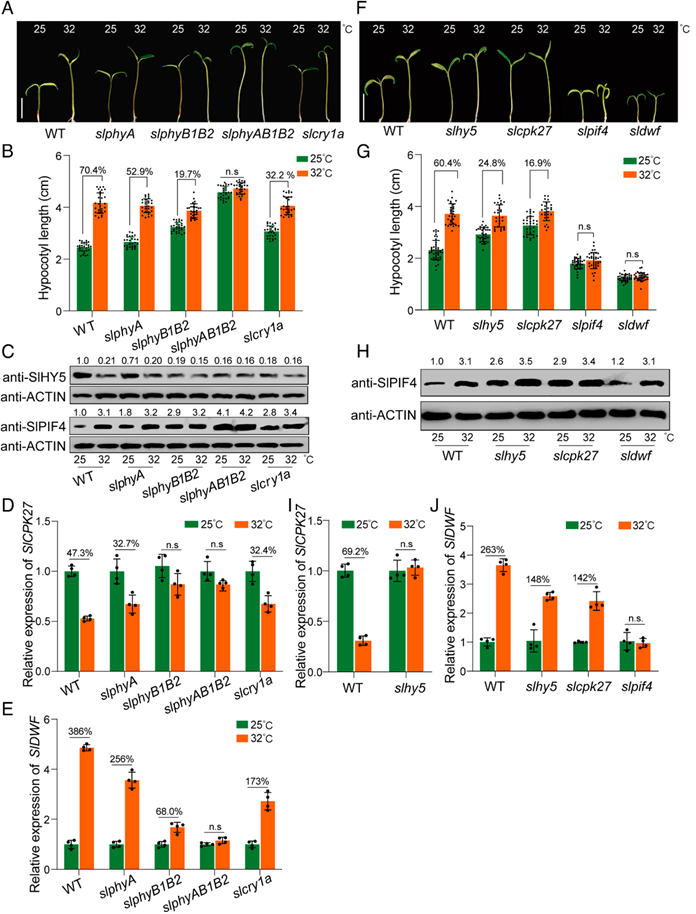 图8 SlHY5-SlCPK27-SlPIF4-BR模块参与番茄热形态建成
图8 SlHY5-SlCPK27-SlPIF4-BR模块参与番茄热形态建成
(图源:Zhu C,Hu Z,Hu C, et al.,Proc Natl Acad Sci U S A. 2024)
二、文章小结
光形态和热形态是植株萌发初期重要的调控,本文章通过转录组和WB实验探究了Ca2+参与光信号传导,通过下游信号通路SlHY5-SlCPK27-SlPIF4-BR诱导植株下胚轴伸长的过程和相关过程中的调控,如Ca2+流动和磷酸化等。这种多用途信号模块表明,光和温度信号转导可能协同进化,从而赋予植物更大的灵活性,微调其生理过程,以更好地适应环境的动态波动
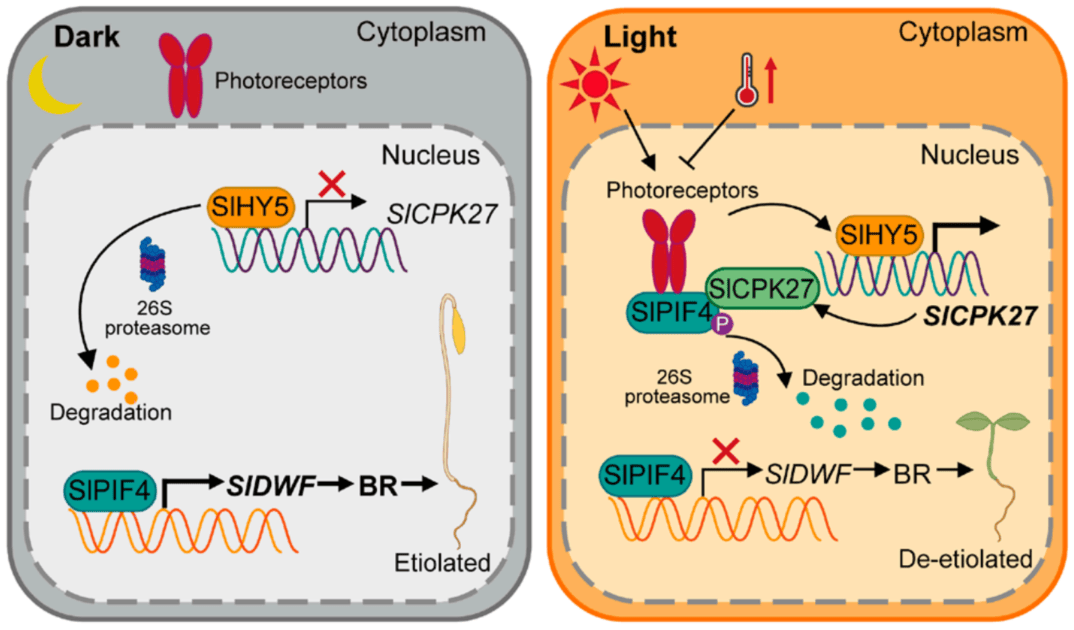 图9 SlHY5、SlCPK27、SlPIF4和BR组成的光信号调控通路模拟图
图9 SlHY5、SlCPK27、SlPIF4和BR组成的光信号调控通路模拟图
(图源:Zhu C,Hu Z,Hu C, et al.,Proc Natl Acad Sci U S A. 2024)
三、拜谱小结
研究采用了突变株/突变株杂交验证,酵母双杂交,原核表达等实验方案,RNA-seq、RT-qPCR、WB、ChIP-qPCR、Co-IP等检测手段,发现SlHY5在光信号传导中增加SlCPK27的转录,激活光信号的传导,SlCPK27通过磷酸化SlPIF4使其降解,SlPIF4通过直接激活SlDWF基因的转录来调控BR的生物合成,从而抑制了下胚轴的伸长。拜谱生物作为一家国内领先的多组学服务公司,可提供完善成熟的蛋白质组学、修饰蛋白质组学(如超高深度磷酸化修饰)、代谢组学、转录组学等多组学产品技术服务体系,整合多组学数据进行深入挖掘分析,全面解析机制机理等,助力高分文章发表。欢迎咨询!
参考文献:
Zhu C, Hu Z, Hu C, Ma H, Zhou J, Xia X, Shi K, Foyer CH, Yu J, Zhou Y. SlCPK27 cross-links SlHY5 and SlPIF4 in brassinosteroid-dependent photo- and thermo-morphogenesis in tomato. Proc Natl Acad Sci U S A. 2024 Sep 3;121(36):e2403040121. doi: 10.1073/pnas.2403040121 Epub 2024 Aug 27. PMID: 39190354;PMCID: PMC11388283





关注公众号





关注小红书


